
Être endurant, c’est aussi ‘En-Durer’, c’est-à-dire résister à la fatigue. Un livre comme celui-là ne suffirait pas pour expliquer la fatigue, phénomène complexe s’il en est. Il existe surtout beaucoup de croyances autour de la fatigue. On reviendra sur le rôle de l’acide lactique sur les crampes et sur les courbatures un peu plus tard. Mais même lors d’efforts intenses, il a été montré récemment que l’acide lactique ne serait pas le ‘serial killer’ accusé de tous les maux. Encore plus étonnant : une étude publiée dans une très sérieuse revue scientifique avait comme titre : Protective effects of lactic acid on force production in rat skeletal muscle (Nielsen et al. 2001). Pas besoin de comprendre l’anglais, vous avez bien lu : l’acide lactique pourrait protéger contre la fatigue ! Pourtant, il existe de nombreuses études montrant que l’ajout de substances pour diminuer l’acidose avait un effet positif sur la performance, donc on ne peut complètement innocenter cette dernière. Elle joue sans conteste un rôle délétère sur la performance. Par quels mécanismes ? Il se peut que son rôle soit indirect, c’est-à-dire via les fibres nerveuses sensitives (‘les cuisses qui brûlent’) qui vont à leur tour limiter la commande centrale ou augmenter l’envie de ralentir ou de s’arrêter. En d’autres termes, l’acide lactique toucherait plus le système nerveux que le muscle.
Il existe aussi pas mal d’idées fausses sur la fatigue en ultra-trail, au premier rang desquelles ce sacro-saint acide lactique. On a vu que son rôle n’était peut-être pas aussi évident, y compris dans les efforts dits ‘lactiques’ justement. Alors en ultra, pensez-vous ! Votre taux de lactate reste au ras des pâquerettes. Sauf bien sûr si vous prenez le départ nettement au-dessus de votre niveau et si, de surcroît, ce surrégime est combiné à un stress important vous faisant produire des catécholamines. Celles-ci vont en effet dégrader plus de glycogène qu’à l’accoutumée, former du pyruvate et donc du lactate en pagaille.
Nous reviendrons, au début du chapitre 3 consacré à la santé, sur les conséquences physiologiques globales d’un ultra-trail. Mais voyons tout de suite ce qu’il en est de la fatigue. Nos nombreuses études conduites à l’université et au CHU de Saint-Etienne et visant à étudier la fatigue extrême en laboratoire (24h sur tapis roulant) et sur le terrain (UTMB® 2009, 2012, 2019 notamment) permettent de répondre en partie à cette question. Ce qui est étudié ici, ce n’est pas seulement le fonctionnement du muscle lui-même, mais de la fonction neuromusculaire dans son ensemble, c’est-à-dire les pertes de force totales (voir Figures ci-dessous : pertes de force et récupération après l’UTMB® et récupération après la WS100) avec ses deux composantes : centrale (ou nerveuse) et périphérique (ou musculaire). Etant entendu qu’il existe une interaction très fine entre ces deux organes, le muscle étant sous la dépendance directe du cerveau, lui-même renseigné en permanence sur le fonctionnement musculaire. Pour en savoir plus sur les origines potentielles de la fatigue, c’est ici.
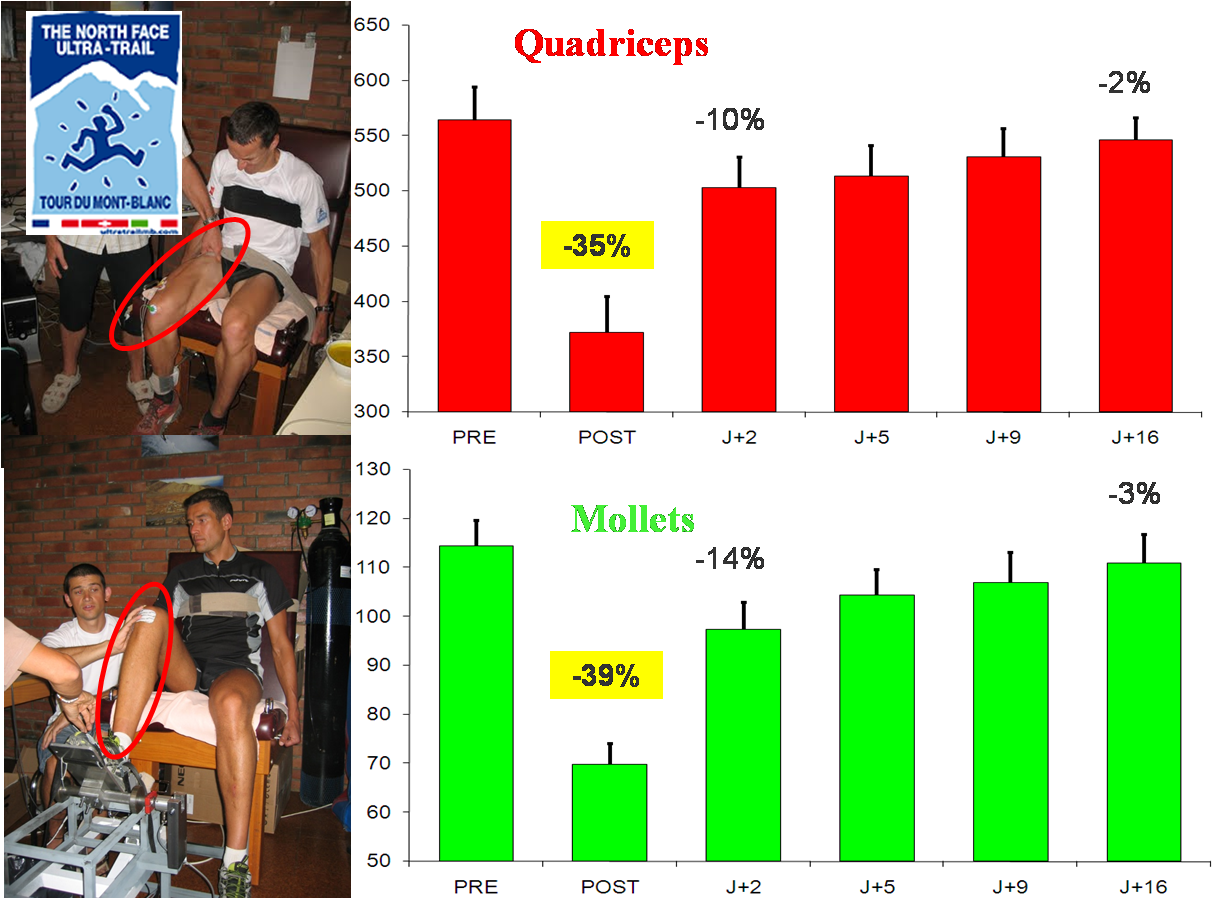
Pertes de force (indice de fatigue neuromusculaire globale) après l’UTMB® et récupération 2, 5, 9 et 16 jours après la course pour les quadriceps (en haut) et les mollets (en bas). D’après Millet et al. 2011
Commençons par le muscle : -10%, c’est la valeur moyenne de perte de contractilité du quadriceps après 24 h de course sur tapis roulant. C’est peu non ? Cette valeur moyenne cache en réalité une très forte variabilité, sans qu’aucune raison évidente (niveau d’entraînement, niveau de performance, expérience, âge) ne puisse expliquer ces larges différences entre les coureurs. Certains semblent donc solides sur le plan musculaire comme d’autres sont gros, grands ou possèdent une grosse VO2max. Ceci peut expliquer entre parenthèses expliquer les incroyables enchaînements de compétitions réalisés avec succès par quelques traileurs(euses) comme François D’Haene en 2019 (doublé Hard Rock-UTMB®). Moins 10% pour le quadriceps donc en moyenne après 24 h de course à plat. On peut imaginer que ce muscle souffre plus en descente : bizarrement, et même si les méthodes n’étaient pas tout à fait les mêmes, il semble que les résultats ne soient pas si différents après un UTMB®, c’est-à-dire pertes assez modérées de contractilité et grosse variabilité entre les coureurs, que ce soit au niveau de la perte de force musculaire ou des indices sanguins de la souffrance musculaire. Deuxième surprise, par rapport à l’étude 24 h tapis, c’est le mollet qui déguste le plus, sans que l’on sache s’il s’agit des effets prioritaires de la montée ou des contractions excentriques en descente. Les deux études, et les nouvelles réalisées depuis, se rejoignent par contre sur le fait que les plus grosses altérations pour le quadriceps ne se situent pas au niveau du muscle lui-même mais de la commande qui provient du cerveau. Là les baisses sont spectaculaires, de -20% à -30% !
En réalité, les choses sont un peu plus compliquées. L’ordre de se contracter qui est envoyé par le cortex moteur est modulé par des informations en provenance de divers récepteurs périphériques dans les muscles, tendons ou articulations. Il faut donc garder à l’esprit que si la fatigue centrale s’installe, et c’est le cas comme le montre la baisse de l’activation maximale du muscle, ce n’est pas le cerveau seul qui est responsable mais de façon plus générale le système nerveux. Pour expliquer cette large fatigue centrale, nous privilégions la piste des récepteurs à l’étirement qui se trouvent dans des petites fibres parallèles aux fibres musculaires plus classiques (Millet et al., 2018). Soyons honnêtes, nous n’avons pas pu apporter la preuve absolue cette origine de la fatigue centrale, il s’agit plutôt là d’une conclusion par défaut.
Au total, quand on compile la fatigue du quadriceps pour toutes les études de course à pied entre 30 min et 35 h, on obtient la figure e-1.2 : augmentation non-linéaire qui tend à un plateau. Comme si les sujets refusaient en moyenne de perdre plus de 40%. Cela peut paraître beaucoup mais de nombreux exercices plus courts (notamment excentriques) induisent des niveaux de fatigue plus élevés et des récupérations plus longues. En réalité, dans une étude sur le Tor des Géants, la perte de force des quadriceps n’était que de 25% (Saugy et al., 2013). Conclusion : il est moins fatiguant de courir 200 miles que 100 (Giandolini et al., 2016)… On voit bien là que la fatigue neuromusculaire n’explique pas tout.
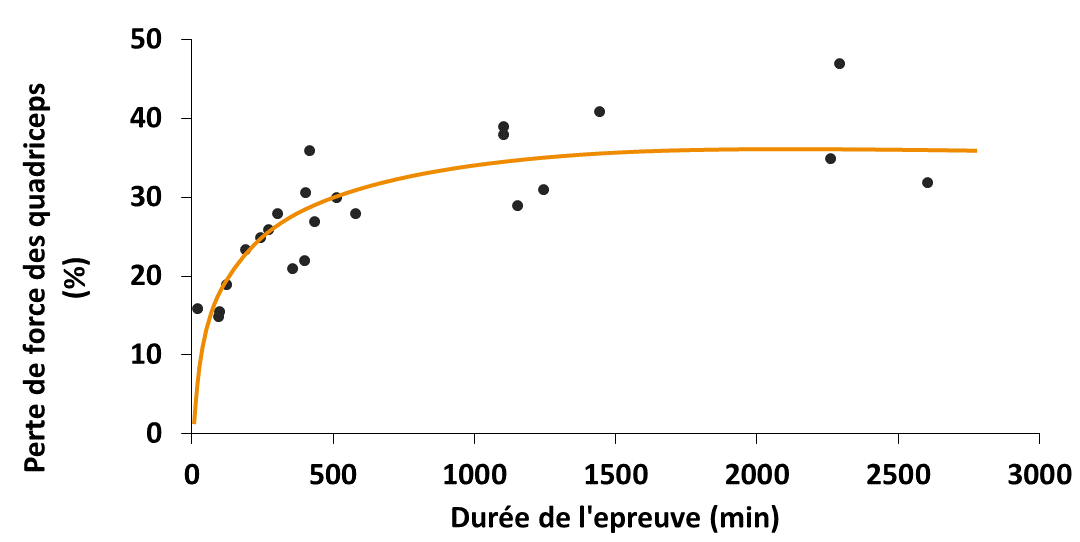
Evolution de la fatigue au niveau des quadriceps pour des épreuves de course à pied de durée variable. Adapté de Millet 2011
Muscles respiratoires et équilibre : les autres effets de la fatigue
Dans les disciplines plus intenses, les muscles respiratoires, qui peuvent se fatiguer comme les autres, représentent parfois une limite à la performance. A tel point qu’il existe des appareils pour les entraîner, soit en force (type Powerbreath®), soit en endurance (type SpiroTiger®). Toutefois, cet investissement ne semble pas utile au coureur d’ultra, la fatigue des muscles respiratoires n’étant vraisemblablement pas un facteur limitant sur la longue distance.
Par Phanélie Berthon, Université de Savoie
Comme vous le savez sûrement, les principaux nutriments énergétiques disponibles pour faire fonctionner vos cellules (dont vos cellules musculaires) sont les glucides et les lipides aussi bien au repos qu’à l’exercice. Les lipides sont des nutriments essentiels pour maintenir le corps en bonne santé. Ils jouent un rôle majeur comme susbtrats énergétiques, sont les principaux composants des membranes cellulaires ou encore permettent l’absorption intestinale des vitamines liposolubles.
Mais commençons par le début ! Au repos, les cellules du corps utilisent en moyenne 50 à 60% de substrats lipidiques et 40 à 50% de substrats glucidiques pour fonctionner. Attention, nous sommes tous différents et ces chiffres peuvent varier légèrement d’une personne à l’autre, en fonction du niveau d’entraînement (voir partie ci-dessous) ou encore du régime alimentaire (voir régime cetogène).
Avant d’aller plus loin, il est utile de savoir d’où viennent, et comment l’organisme stocke, les lipides et les glucides utilisés par les muscles à l’effort. C’est notre alimentation qui fournit au corps les substrats énergétiques glucidiques et lipidiques. Une fois digéré, les lipides (ou acides gras libres = AGL) et les glucides (principalement le glucose) traversent la paroi intestinale et arrivent dans le sang. On parle alors de glucose sanguin et de lipides ou d’AGL sanguins. 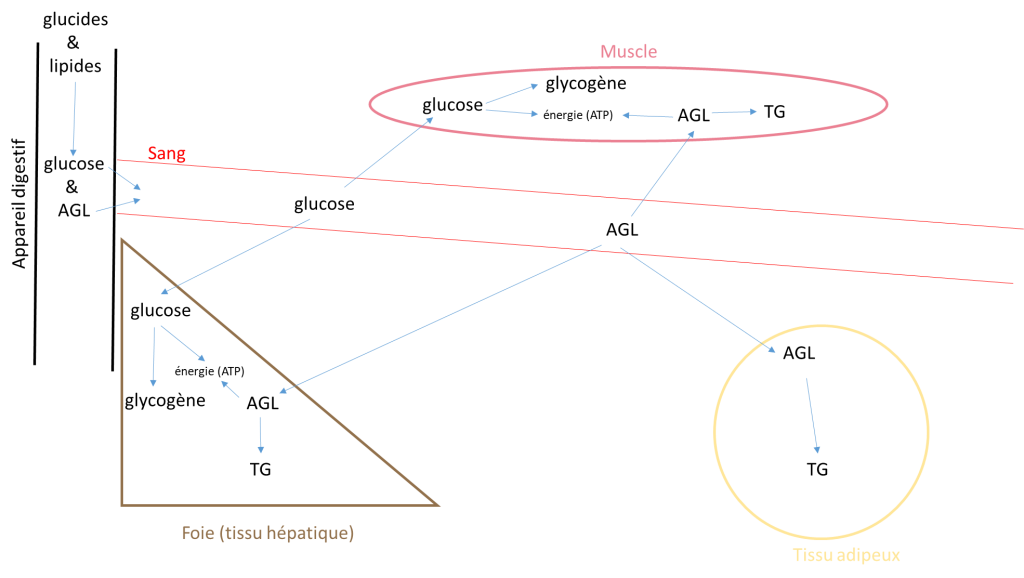 Ces substrats circulants (glucose et AGL) peuvent être soit utilisés immédiatement par les cellules du corps qui les transforment en énergie soit stockés. Le glucose est stocké seulement dans les muscles et le foie sous forme de glycogène (grosses molécules obtenues par des glucoses fixés les uns aux autres) et les lipides sont stockés majoritairement dans le tissu adipeux, et dans une moindre mesure dans les muscles sous forme de triglycérides ou TG (3 AGL fixés ensembles).
Ces substrats circulants (glucose et AGL) peuvent être soit utilisés immédiatement par les cellules du corps qui les transforment en énergie soit stockés. Le glucose est stocké seulement dans les muscles et le foie sous forme de glycogène (grosses molécules obtenues par des glucoses fixés les uns aux autres) et les lipides sont stockés majoritairement dans le tissu adipeux, et dans une moindre mesure dans les muscles sous forme de triglycérides ou TG (3 AGL fixés ensembles).
Que se passe-t-il à l’exercice ? Chaque exercice est différent en fonction de son intensité et sa durée et cela va influencer le fonctionnement des muscles et donc l’utilisation des substrats énergétiques. Une étude, assez ancienne mais qui est une référence dans le domaine (Romijn, 1993) nous le montre clairement.
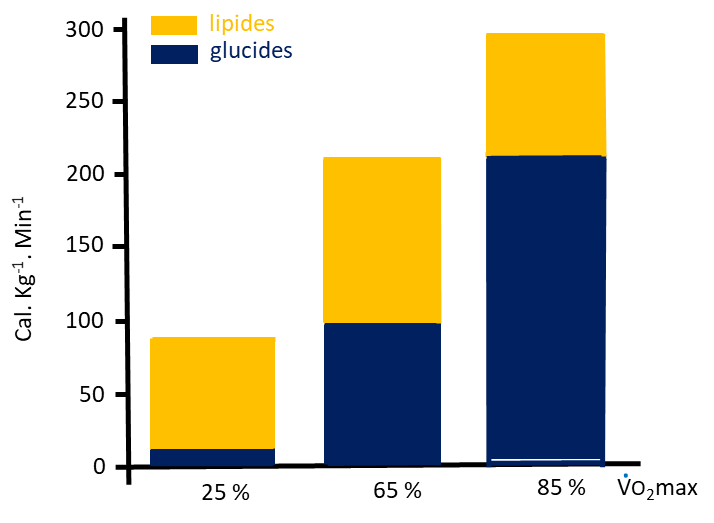
Cette première figure présente, pour 3 intensités d’exercices différentes (25, 65 et 85% de VO2max), la quantité d’énergie libérée par la dégradation des lipides (AGL circulants et triglycérides musculaires) et des glucides (glucose circulants et glycogène musculaire). Comme vous pouvez le voir, ce sont pour les faibles intensités d’exercice que nous utilisons proportionnellement le plus de lipides et à l’inverse, les glucides sont les principaux substrats quand l’intensité de l’exercice est élevée. Lors d’un ultratrail, l’intensité moyenne est de 40 à 50% de VO2max, et donc les lipides sont des substrats énergétiques non négligeables dans ce type d’exercice !
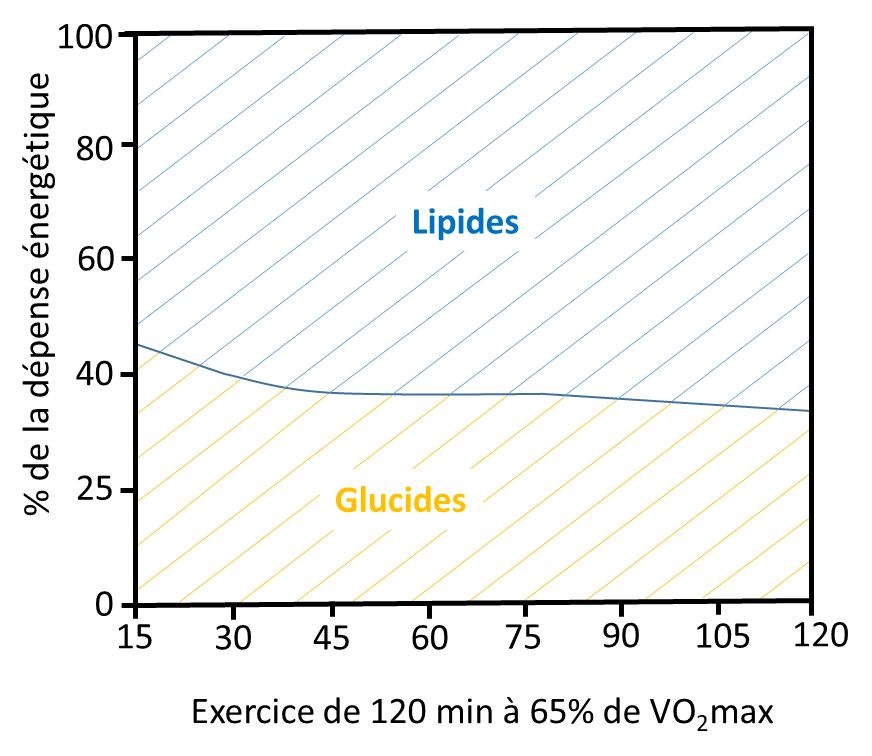
Cette première figure présente, pour 3 intensités d’exercices différentes (25, 65 et 85% de VO2max), la quantité d’énergie libérée par la dégradation des lipides (AGL circulants et triglycérides musculaires) et des glucides (glucose circulants et glycogène musculaire). Comme vous pouvez le voir, ce sont pour les faibles intensités d’exercice que nous utilisons proportionnellement le plus de lipides et à l’inverse, les glucides sont les principaux substrats quand l’intensité de l’exercice est élevée. Lors d’un ultratrail, l’intensité moyenne est de 40 à 50% de VO2max, et donc les lipides sont des substrats énergétiques non négligeables dans ce type d’exercice !
Si nous nous intéressons maintenant à la durée de l’exercice, il apparait très clairement que pour un exercice sous maximal à 65% de VO2max, la part des lipides utilisés comme substrats énergétiques augmente avec la durée de l’exercice. Dans cette étude, l’exercice ne durait que 2 h, on est loin de la durée d’un ultratrail ! D’autres travaux ont été réalisées en laboratoire sur un exercice de 24h sur tapis roulant et des résultats similaires ont été retrouvés, avec une augmentation progressive de la part des lipides comme substrat énergétique, au moins sur les premières heures de l’exercice.
Evolution du quotient respiratoire sur une épreuve de 24 h sur
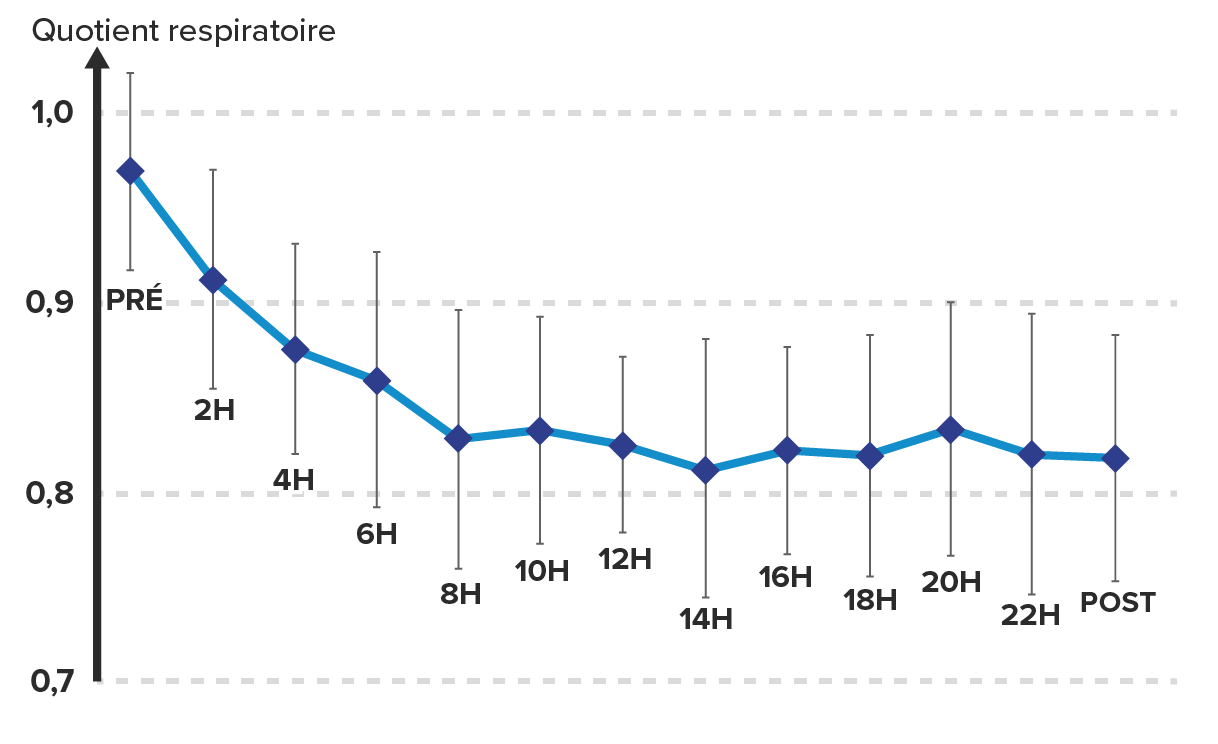
tapis roulant. D’après Gimenez 2013.
Evidemment, on ne se lance pas dans un ultratrail sans être à minima entraîné, or l’entraînement va également modifier un peu l’utilisation des substrats énergétiques. Il va permettre d’augmenter la part relative des lipides utilisés (pour une même intensité d’exercice) pour répondre à la demande énergétique.
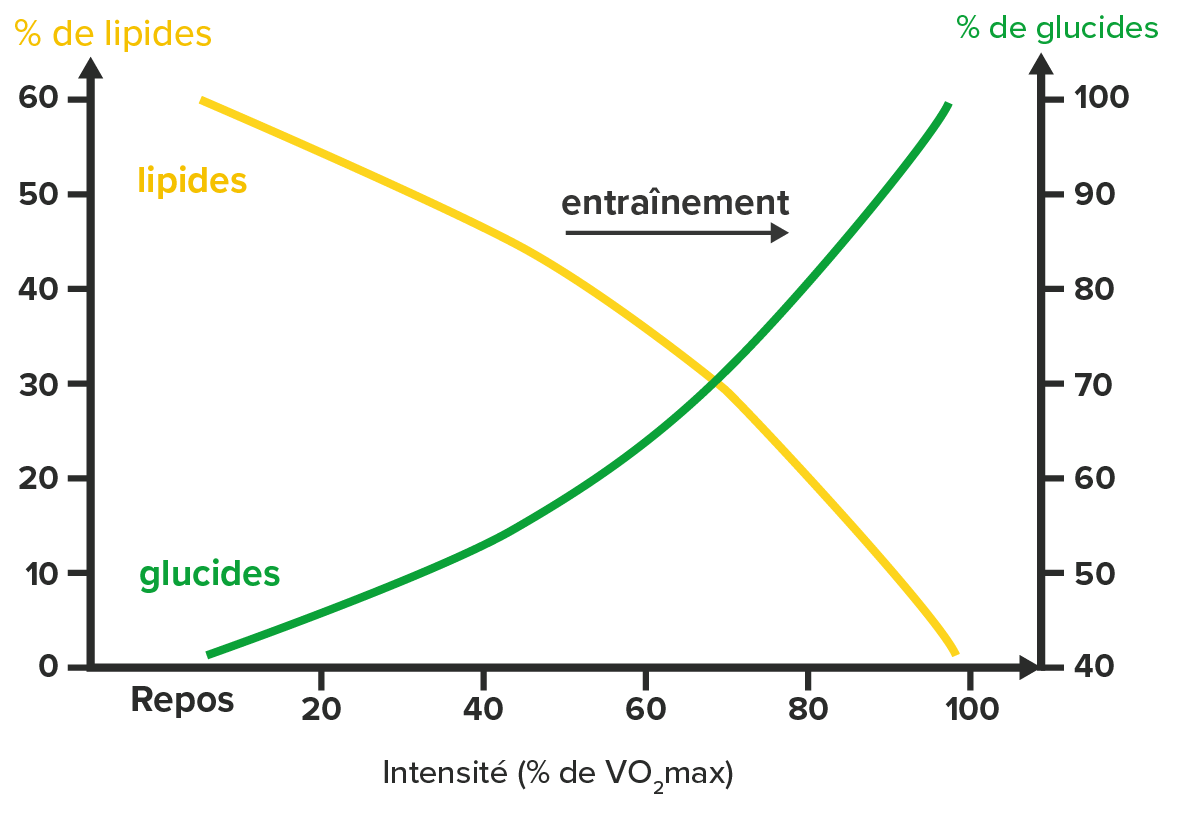
Mais focalisons-nous sur les lipides et essayons de comprendre comment leur utilisation peut varier en fonction de l’intensité, de la durée de l’exercice mais aussi de l’entraînement.
Pour comprendre ces changements cellulaires, commençons par décrire les 4 étapes qui permettent aux lipides d’être utilisés comme substrat énergétique par les cellules. Les lipides sont majoritairement stockés dans le tissu adipeux (sous la peau et autour des viscères), c’est-à-dire « loin » du muscle mais aussi dans les muscles, même si les quantités sont beaucoup plus faible.
La première étape est donc la mobilisation et le transport des AGL du tissu adipeux vers les muscles grâce à la circulation sanguine. Une étude de 2007 a très bien mis en évidence une disponibilité accrue des acides gras dans le sang lors d’une épreuve d’ultra-endurance de 411 km en environ 100 heures (Helge, 2007).
La seconde étape est leur entrée dans les cellules musculaires. Certains AGL (les plus petits) vont pouvoir traverser la membrane cellulaire par diffusion, mais ce mécanisme est minoritaire. La plupart des AGL vont devoir utiliser un processus hautement régulé faisant intervenir des protéines de transport spécifiques aux AGL, protéines qui sont enchâssées dans la membrane cellulaire. La plus connue de ces protéines porte un nom bien peu attrayant : FAT/CD36. Une étude sur des animaux génétiquement modifiés (suppression de la protéine membranaire FAT/CD36) montrent très clairement que l’absence de FAT/CD36 est liée à une baisse importante de l’entrée des lipides dans les cellules musculaires et ceci est associé à une moindre utilisation des lipides comme substrat énergétique, et donc à une plus grande utilisation des glucides (McFarlan, 2012). A l’inverse, plus il y a de protéines de transport dans la membrane, plus la quantité d’AGL qui pénètre dans la cellule est importante. Cela sous-entend que les cellules musculaires sont capables d’adapter la quantité de FAT/CD36 dans leur membrane. Pour cela différents mécanismes sont décrits :
1-A l’intérieur des cellules musculaires, il y a des stocks de protéines FAT/CD36 qui vont pouvoir être mobilisées (c’est-à-dire se déplacer vers la membrane cellulaire) en quelques minutes. Ce mécanisme s’appelle la translocation (Bonen, 2000). Cela survient quand un muscle se contracte, donc quand on fait un exercice. Ce mécanisme chez l’homme est difficile à étudier car cela nécessite des prélèvements de muscles assez conséquents. Heureusement, les chiens de traineaux sont là ! Une étude sur des chiens entraînés pour des courses sur de très longues distances comme La Grande Odyssée a montré, en réponse à un exercice (25 min), il y avait une augmentation de l’entrée des lipides dans les muscles associés à une augmentation de la translocation des protéines FAT/CD36. Mais, assez étonnamment, ce mécanisme n’est pas amplifié si les chiens ont fait 4 jours d’exercice successif (simulation de course) (Davis, 2014).
2- Quand ces exercices d’endurance se répètent (c’est ce que l’on appelle un entraînement !) la quantité totale de protéines FAT/CD36 va augmenter (via l’augmentation de l’expression des gènes codant pour cette protéine) en quelques jours (Tunstall, 2002).
Les mêmes mécanismes existent d’ailleurs pour le transport du glucose dans les cellules musculaires, avec les protéines de transport GLUT4.
La troisième étape est leur transport vers la structure intracellulaire capable de les dégrader pour libérer de l’énergie, la mitochondrie. La mitochondrie, si vous préférez, est l’usine qui permet de produire de l’énergie dans les cellules. Les lipides vont donc devoir « entrer » dans ces usines, et la protéine de transport FAT/CD 36 va encore une fois intervenir au niveau de la membrane des mitochondries (Bonen, 2000). Dans une étude que nous avons citée plus haut, ils montrent également l’augmentation de l’utilisation musculaire des acides gras (Helge, 2007).
Et nous voici enfin à la quatrième et dernière étape : Les lipides vont être transformés en énergie utilisable par les muscles ! Pour cela, ils vont être dégradés grâce à un autre type de protéines, les enzymes. La cellule est capable d’augmenter l’efficacité de certaines de ces enzymes, ou si vous préférez leur rendement au travail. C’est ce qui se passe lors d’un exercice à condition qu’il ne se prolonge pas trop longtemps. En effet il semblerait que pour des exercices de très longue durée (24h et plus), l’efficacité des enzymes produisant l’énergie dans la mitochondrie soit altérée (Fernstrom, 2007). Mais grâce à l’entraînement, vous pourrez augmenter l’efficacité de travail de ces enzymes, et stimuler la synthèse de nouvelles protéines (stimulation de l’expression des gènes). Il y aura donc plus d’enzymes capables de dégrader les lipides pour produire de l’énergie !
Pour conclure, les lipides ne comptent pas pour du beurre chez les ultratraileurs ! Alors, consommez-en, et consommez les bons ! Les recommandations de l’ANSES (Agence Nationale de Sécurité Sanitaire Alimentaire) sont les suivantes : Vous devez avoir un apport en lipides représentant 35- à 40 % des apports nutritionnels conseillés (ANC). Attention à ne pas trop diminuer les apports en lipide sous peine d’être carencé en certains acides gras dits « essentiel » car non synthétisés par le corps. Il s’agit des acides gras oméga-3 et oméga-6. Mais c’est un autre sujet !
Nous avons eu l’opportunité de tester Philippe Fuchs, un ultramarathonien qui a couru de Paris à Pékin pour les J.O. de 2008, soit 8 500 km en 161 jours (Millet et al., 2009). Il a été testé sur un tapis d’analyse de la foulée avant et après son raid. Comme le montre la figure ci-dessous, sa technique de course s’est grandement modifiée dans le sens d’une augmentation de sa fréquence de la foulée, une diminution du temps passé en l’air entre deux appuis, ce qui induisait une diminution du pic d’impact. Nous avons émis l’hypothèse que cette stratégie était pour lui un moyen de préserver son intégrité physique malgré le fait que cette technique soit plus dispendieuse sur le plan énergétique puisqu’en parallèle, son Cr augmentait de 6,2%.
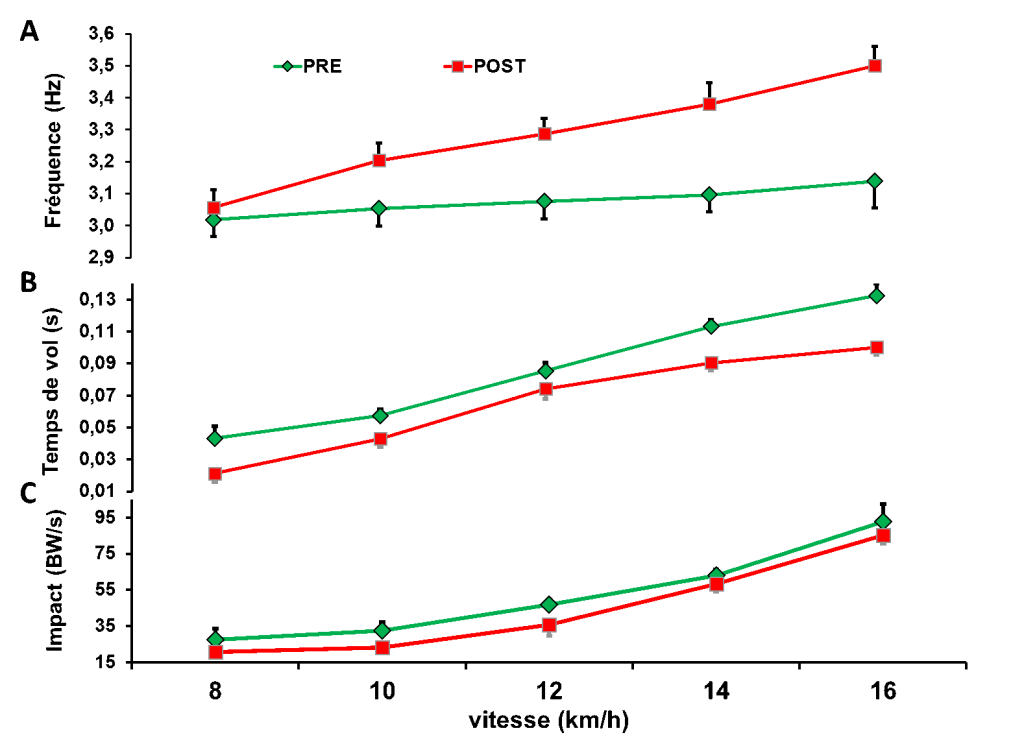
Evolution de la fréquence de foulée (A), du temps de vol entre deux appuis (B) et du pic d’impact au sol (C) à différentes vitesses de course mesurés 3 semaines avant (PRE, vert) et 3 semaines après (POST, rouge) un raid en course à pied de 8 500 km en 161 jours. Adapté de (Millet et al., 2009).
